| † Ихтиостега | |
| Часть скелета в Палеонтологическом музее им. Ю. А. Орлова, Москва | |
| Модель в Государственном музее естествознания Штутгарта | |
| Царство: | Животные |
| Подцарство: | Эуметазои |
| Без ранга: | Двусторонне-симметричные |
| Без ранга: | Вторичноротые |
| Тип: | Хордовые |
| Подтип: | Позвоночные |
| Инфратип: | Челюстноротые |
| Клада: | Тетраподоморфы[1] |
| Семейство: | † Ichthyostegidae |
| Род: | † Ихтиостега |
Säve-Söderbergh, 1932
| 36965 |
Ихтиосте́га
[2] (лат. Ichthyostega, от др.-греч. ἰχθυο- + στέγη — рыбья кровля) — род вымерших челюстноротых из семейства Ichthyostegidae клады тетраподоморф (Tetrapodomorpha), живших в девонском периоде[3] и представляющий собой первое промежуточное звено между рыбами и амфибиями. Этот род не является прямым предком современных видов, предки которых — лепоспондилы (Lepospondyli) — появились в каменноугольном периоде.
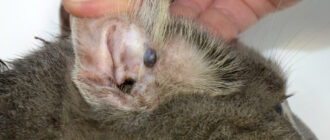
У ихтиостег были лапы, но они, возможно, не использовались для ходьбы по суше. Ихтиостеги имели хвостовой плавник и некоторые органы чувств, функционирующие только в воде. Тело их было покрыто мелкими чешуйками.
По мнению некоторых учёных, ихтиостеги могут считаться переходными формами между кистепёрыми рыбами и наземными позвоночными.
История изучения
В 1932 году Сёве-Седерберг описал четыре вида ихтиостег из верхнего девона, найденных в восточной части Гренландии. Эти виды могли быть синонимичными, потому что их морфологические различия не были резко выражены. Виды различались в пропорциях и строении черепа. Сравнение было проведено на четырнадцати экземплярах, собранных в 1931 году Датской Восточной Гренландской экспедицией. Дополнительные экземпляры были собраны между 1933 и 1955 годами.
Род ихтиостег находится в родстве с акантостегой (Acanthostega gunnari
), также обнаруженной в восточной Гренландии. По сравнению с акантостегой череп ихтиостеги выглядит более рыбообразным, однако её пояс передних конечностей сильнее и лучше адаптирован к передвижению.
Теория эволюции[править | править код]
Ко времени написания Чарльзом Дарвином «Происхождения видов» науке не были известны переходные формы жизни. Этот факт Дарвин считал одним из самых слабых мест своей теории. Однако уже через два года был обнаружен археоптерикс — предположительное переходное звено между рептилиями и птицами.
Переходные формы между двумя группами организмов не обязательно являются потомками одной группы и предками другой. По ископаемым останкам, как правило, невозможно достоверно установить, является ли один организм прямым предком другого. Кроме того, вероятность найти в палеонтологической летописи чьего-нибудь прямого предка очень мала, и по умолчанию среди биологов принято такую возможность вообще не рассматривать. Гораздо выше вероятность обнаружить относительно близких родственников этого предка, схожих с ним по строению. Поэтому любая переходная форма автоматически интерпретируется как боковая ветвь эволюции, а не «участок ствола».[2]
Описание
Ихтиостеги были около 1,5 метров длиной и имели по семь пальцев на задних лапах. Точное количество пальцев на передних лапах пока не установлено, но вероятно, что их было тоже семь. На хвосте у них был плавник «рыбьего» типа, поддерживаемый невральными и гемальными дугами.
Ноздри располагались у нижнего края челюстей. Слёзная кость примыкает к ноздре, но не к глазнице. Межвисочная кость отсутствует (это обстоятельство исключает возможность происхождения большинства темноспондилов от этого рода). Заднетеменная кость непарная. Челюстная кость соприкасается с квадратноскуловой. Сохраняются подкрышечные и предкрышечные кости. Носовые кости широкие. Глазницы овальные и располагаются в центральной части черепа. Хорда через отико-окципитальную часть мозговой коробки доходит до гипофизарной ямы.
Переходные формы. Найдены ли они?
Вступление
Что такое переходные формы? Это виды, в строении которых возникают новые элементы, характерные для их эволюционных потомков, но еще сохраняются старые элементы, доставшиеся в наследство от эволюционных предков. А еще это источник мифов о Теории эволюции, стабильно работающий уже полтора столетия.
Ученые из года в год ищут «переходное звено» от обезьяны к человеку и никак не могут найти. Противники эволюционизма утверждают, что переходных форм нет, а этот ваш археоптерикс – тупиковая ветвь. Журналисты пугают общественность тем, что у человека и банана 50% общей генетической информации. Если птицы и крокодилы родня, то где окаменелости мезозойской крокоутки? – вопрошают диванные скептики. Да и от самого термина несет какой-то недоделанностью. Что это за вид такой, который «ни то ни сё»?
Давайте разбираться.

Древо жизни
Для того чтобы получить представление о переходных формах, надо обратиться к биологической систематике. Идея «разложить по полочкам» все разнообразие живых организмов не нова. В XVIII веке очередной способ классификации был предложен шведским натуралистом Карлом Линнеем. Способ оказался настолько удачным, что после важного апгрейда в середине XIX века используется до сих пор.
В чем тут суть? Неожиданно все формы жизни устроены не абы как. Анатомические структуры – все эти органы и ткани – не разбросаны в хаотичном порядке, а подчиняются определенным закономерностям. Вот, к примеру, позвоночник. Он есть у рыб, птиц, рептилий, зверей и амфибий. Но ни одного жука с позвоночником вы не найдете. Как нет и птиц без позвоночника. А те, у кого есть позвоночник, делятся на рыб (с жабрами и без лап) и четвероногих (с легкими и лапами). А четвероногие, например, могут быть пернатыми, шерстистыми, чешуйчатыми или с голой слизистой кожей. Описание, конечно же, очень упрощенное, но принцип понятен.
Анатомические структуры выстраиваются в иерархическом порядке, от общего к частному, что позволяет разделить все организмы на соответствующие иерархические группы. Отсюда получаем биологическую систематику с известными всем типами, классами, отрядами, семействами, родами и видами.
Но почему живые организмы устроены именно так? Ключом к ответу на этот вопрос стала теория эволюции, предложенная Чарльзом Дарвином в 1859 году. Иерархия анатомических структур отражает ход эволюции, а их единство у разных организмов говорит об общем происхождении. К настоящему времени этот тезис многократно подтвержден фактами из сравнительной анатомии, эмбриологии, палеонтологии и генетики.
Систему биологической классификации можно представить в виде дерева (скорее куста), на котором каждый отдельный листик – это вид, а ветви и стволы – разные таксоны. Такое «дерево» называют филогенетическим, эволюционным, или же Древом жизни.
Из вышесказанного для нас важными будут шесть следствий.

Следствие первое: строение организмов можно предсказать
Могут ли у кошек быть копыта? Если кит плавает в воде, то почему у него нет жабр? Где искать цветок папоротника? Чтобы ответить на подобные вопросы, вовсе не нужно осматривать всех китов и кошек, и следить за всеми папоротниками в Купальскую ночь. Зная биологическую систематику можно заранее сказать, какие анатомические признаки должны присутствовать у той или иной группы организмов.
Такой инструмент очень удобен в палеонтологии. Имея всего один зуб можно классифицировать животное, иногда с точностью до рода. И на основании этого сделать вполне надежную реконструкцию. А уже потом, когда (и если) будет найден полный скелет, можно уточнять детали и делать нашу реконструкцию все более реалистичной.
Но можно обойтись и вовсе без останков. Например, если нас интересует общий предок всех кошачьих, мы можем составить абстрактный портрет, исходя из признаков, характерных для этого семейства. Получается, что окаменелостей нет, а мы уже знаем, кого искать.

Следствие второе: общих родственников можно найти
Древо жизни охватывает абсолютно все виды организмов. А значит мы все друг другу родственники. Можно взять два абсолютно любых вида, а затем спуститься вниз по веточкам нашего Древа, до точки пересечения. В этой точке будет находиться общий предок, а в его строении будет совмещены признаки, общие для обеих ветвей. Даже если идти придется очень далеко вниз.
Например, предок человека и банана по совместительству будет предком вообще всех растений и животных. Это одноклеточный организм, живший как минимум 3 млрд лет назад, то есть раньше самых древних следов растений в горных породах. И наш геном действительно хранит информацию, оставшуюся от этого далекого-далекого предка.

Следствие третье: переходные формы повсюду
От древних одноклеточных организмов до человека – очень долгий путь. Поколение за поколением, вид за видом наши предки менялись под действием эволюции. Древо жизни росло, давало множество новых побегов и в итоге получилось современное видовое разнообразие. Можно ли сказать, что на этом эволюция остановилась? Конечно, нет. Те современные виды, которые не вымрут, дадут свое эволюционное потомство. И человек тут не исключение.
А значит, каждый вид в любой момент времени является потенциально переходным между какими-то старыми и новыми видами. Мы все – переходное звено.
Но не стоит ждать, что наши потомки будут так уж сильно от нас отличаться. Эволюционные изменения накапливаются постепенно, дети все так же похожи на родителей, и заметить отличия можно изменив масштаб. На промежутке в 1 млн лет мы точно сможем различить два вида, но вот найти точку, в которой один вид стал другим – не получится.

Следствие четвертое: некоторые переходные формы интереснее других
Как и в истории, эволюционные события могут иметь разный масштаб. Например, процесс эволюции уток, поиск прото-утки и переходных форм между ними – это все очень интересно. Но на масштабные эволюционные события не тянет. Другое дело – происхождение птиц вообще.
Из современных животных ближайшая родня уток – это крокодилы. Спускаемся по веточкам нашего дерева, находим точку пересечения. Здесь, в этой точке, в триасовом периоде жил их общий предок. Внешне был похож на тощего голенастого крокодила. Имел четырехкамерное сердце, высокий метаболизм и откладывал яйца. Не то чтобы сильно похоже на птицу, но уже кое-что. Значит, в качестве переходной формы нам надо искать кого-то похожего, но с крыльями и перьями. В XIX веке про родство птиц с крокодилами не знали, искали просто пернатого птице-ящера. И нашли. Им оказался археоптерикс, вошедший во все учебники биологии как эталон переходной формы. Так вот он – родоначальник птиц? Вообще-то нет.

Следствие пятое: неуловимые предки
Вообще-то обнаружение переходной формы еще не обозначает обнаружение того самого вида-предка, который дал всю дальнейшую эволюционную ветвь. И чем шире область поиска, чем более древняя у нас переходная форма, тем меньше для этого шансов. Как так?
Переходная форма – это все равно, что фоторобот, набор анатомических структур, которые точно должны быть. Например: скелет рептилии (включая длинный хвост и зубы), легкие кости, перья, небольшой размер. Это сокращает поиск до одной из групп динозавров, живших в юрском периоде. А дальше начинаются проблемы.
В группе – сотни видов, не имеющих узкой специализации. Они жили на промежутке в несколько десятков млн лет, приходились друг другу какими-то родственниками и все обладают тем набором признаков, который нам нужен. То есть потенциально они все – переходные формы к птицам. Но какой шанс, что нам улыбнулась удача и из нескольких сотен видов мы нашли «того самого»?
Разумеется, изучая этих оперенных то-ли еще динозавров, то-ли уже птиц, мы будем накапливать информацию и отсеивать ненужные варианты. Археоптерикс окажется хоть и ранней птицей, но параллельной ветвью эволюции. Наш портрет птицы-первопредка будет становиться все точнее и точнее. Можно будет локализовать поиск до ранней юры Северного Китая, а таксономию до манирапторов. Но указать на конкретный вид-предок мы вряд ли сможем.

Следствие шестое: и все-таки их можно отыскать
Так ли уж важно знать, какой из десятка видов птицеподобных динозавров был точно первоптицей? Для специалистов важно, для широкого круга людей – пожалуй, не очень. Что уж говорить о предках жесткокрылых или голосеменных растений. А если о происхождении человека?
Про нас самих нам нужно знать как можно более точно. И в этом плане в сравнении с птицами нам крупно повезло. Все-таки мы изучаем молодые останки, большинству которых нет и 1 млн лет. Их достаточно много, сохранность хорошая, а если они моложе 100 тыс лет – есть возможность использовать палеогенетику. А значит, мы можем не просто указать на переходную форму от австралопитеков к нам, но и восстановить нашу эволюцию, шаг за шагом.
Сегодня вместо давно устаревшей линейной схемы, мы имеем разветвленное деревце, объединяющее наших родственников и предков. Это все равно что семейная фотография с несколькими поколениями. Мы с большой точностью можем назвать своих родителей и их родителей. А вот с остальными не всегда понятно, кто с кем в каком родстве состоит. Антропология развивается стремительно, создаются новые методы исследований и делаются замечательные открытия. Разумеется, восстановить всю цепочку поколений (как того иногда требуют креационисты) мы не сможем. Но серьезно увеличить детальность и сложность и без того непростой картины антропогенеза ученым вполне по силам.

Заключение
Подведем итог.
Переходные формы действительно существуют. И в большинстве своем они объединяют группу родственных видов, имеющих специфический набор анатомических особенностей. Часть этих особенностей новые и они получат развитие у потомков. Часть – старые, доставшиеся от предков и к потомкам они не перейдут. Особый интерес представляют переходные формы между крупными таксонами. Например, между рыбами и амфибиями. Или рептилиями и птицами, рептилиями и млекопитающими. На их примере можно видеть, как незначительные изменения приводят к серьезным результатам.
Благодаря развитию палеонтологии известно множество переходных форм. Эволюция позвоночных животных, например, изучена достаточно детально. И самая, пожалуй, изученная область – это эволюция человека. И все-таки, белых пятен хватит еще не на одно поколение палеонтологов. А учитывая неполноту геологической летописи, весь паззл мы не соберем никогда. Тем важнее вклад каждого, кто решил связать свою жизнь с палеонтологией. Возможно, и вы станете первооткрывателями недостающего звена.

Примечания
- Из-за ограничений MediaWiki в качестве вышестоящего таксона тетраподоморф указан инфратип челюстноротых (Gnathostomata) вместо клады дипнотетраподоморф (Dipnotetrapodomorpha), приведённого в источнике: Betancur-R. R. et al.
The tree of life and a new classification of bony fishes // PLOS Currents Tree of Life. — 2013. — doi:10.1371/currents.tol.53ba26640df0ccaee75bb165c8c26288. - Ихтиостега // Италия — Кваркуш. — М. : Советская энциклопедия, 1973. — (Большая советская энциклопедия : [в 30 т.] / гл. ред. А. М. Прохоров ; 1969—1978, т. 11).
- ↑ 12Ichthyostega
(англ.) информация на сайте Paleobiology Database. (Проверено 16 мая 2020).
Спор о постепенности эволюционных преобразований[править | править код]
Переходные формы являются объектом исследований в споре между градуалистами и пунктуалистами. Градуалисты считают, что эволюция является постепенным, непрерывным процессом. Пунктуалисты, или сторонники теории прерывистого равновесия считают, что виды в течение долгого времени остаются неизменными, а изменения происходят в очень короткий промежуток времени. Переходные формы являются аргументом обеих сторон. Пунктуалисты утверждают, что большинство видов появляются внезапно и на протяжении всей своей истории меняются слабо. Градуалисты отмечают, что среди позвоночных нет ни одного бесспорного примера очень быстрого перехода от одного вида к другому, большинство переходов вообще плохо задокументированы ископаемыми. Также много хорошо описанных внутривидовых изменений. Например, описаны изменения среднекайнозойских кроликов, миоценовых грызунов, гиппарионов, эоценовых парнокопытных, переход между родами эоценовых приматов.[3]
Примечания[править | править код]
- Доказательства эволюции Палеонтологические доказательства
- Александр Марков.
Ископаемые рыбы в очередной раз подтвердили правоту Дарвина
(неопр.)
(недоступная ссылка). Элементы.ру. Дата обращения 23 ноября 2010. Архивировано 20 августа 2011 года. - Кэрролл Р. Палеонтология и эволюция позвоночных: В 3-х т. Мир, 1992 Т.3 — с. 146
- Эволюция конечностей лошади (неопр.)
(недоступная ссылка). Государственный Дарвиновский музей. Дата обращения 10 июня 2010. Архивировано 26 декабря 2011 года.
Древние амфибии
Глобальный экосистемный кризис, завершивший историю палеозойских сообществ наземных позвоночных, привел к перестройке их главного амфибийного компонента – группы Temnospondyli. К концу перми она была почти вытеснена из наземных биотопов и сохранялась как заметный элемент лишь в виде двух географически изолированных водных семейств – Rhinesuchidae и неотенических Dvinosauridae.
Но события, связанные с консолидацией и воздыманием суши при переходе к триасу, привели к новому повороту в судьбе группы. Рост аридизации и редукция низинных пространств означали сужение зон, пригодных для жизни позвоночных на суше, что дало преимущества обитателям водоемов, не затронутым этими ограничениями. Соответственно, в раннем триасе отмечается взрывной рост разнообразия новых водных темноспондильных семейств. В большинстве они не пережили этого интервала, хотя успели достигнуть (в разной степени) почти всесветного распространения.
Дальнейшая история Temnospondyli связана в основном с их капитозавроидными
линиями, а также с небольшим числом слабо диверсифицированных групп, таких как
брахиопиды, метопозавры, плагиозавры
и т.п., впервые получивших достаточно заметное распространение лишь в среднем или позднем триасе.
(М.А. Шишкин)
Ссылки[править | править код]
- Миф о том, что «недостающее звено не найдено…» на портале Антропогенез.ру.
- Переходные формы: как их нашли и не нашли.
- Соколов А. Б.
Недостающее звено. Мифы об эволюции человека. ScienceVideoLab.
Для улучшения этой статьи желательно:
Пожалуйста, после исправления проблемы исключите её из списка параметров. После устранения всех недостатков этот шаблон может быть удалён любым участником. |
Антракозавры (амфибии, от которых произошли рептилии)
Антракозавры (лат. Anthracosauria), или эмболомеры
(лат. Embolomeri) — группа наиболее примитивных
рептилиоморф
, или
батрахозавров
. Иногда «антракозаврами» называют всех рептилиоморф. Череп относительно высокий, иногда сжатый с боков. Сохраняется кинетизм черепа — как у кистепёрых рыб — подвижность между «щеками» и крышей черепа. Барабанной перепонки, видимо, нет, хотя существует вырезка по заднему краю щеки. Зубы обычно мощные, часто есть «клыки» на небе. Позвоночник эмболомерного типа. Тело удлиненное, хвост мощный, сжатый с боков. У некоторых описан «хвостовой плавник» из удлиненных супраневральных радиалий, как у рыб. Конечности у наземных форм короткие, но мощные, у водных форм — слабые. (Из Википедии)
Филогенетические ряды[править | править код]
Филогенетический ряд лошади. Показана реконструкция ископаемых видов, полученных из последовательных слоёв породы. Справа — вид спереди передней левой ноги с выделенным третьим сегментом, а также зуб в продольном сечении[4].
Филогенетические ряды − ряды видов, последовательно сменявших друг друга в процессе эволюции различных групп животных и растений.
Впервые были открыты В. О. Ковалевским, который показал, что современные однопалые копытные происходят от древних пятипалых мелких всеядных животных.